文、圖表/李璠(行政院農業委員會家畜衛生試驗所疫學研究組研究員兼組長)
家禽流行性感冒病毒(以下簡稱禽流感病毒)是A型流行性感冒病毒的一支,凡是能夠感染鳥類,或從禽鳥檢體檢測到的A型流行性感冒病毒,都稱為「禽流感病毒」。
自從1990年代開始,源自於中國的H5亞型高病原性禽流感病毒逐漸演化成多種具有高致病力與高傳染力的H5亞型禽流感病毒,造成全球數波大流行,對歐洲、亞洲、北美洲的家禽產業的殺傷力極強。
流行的禽流感病毒亞型,包括H5N1、H5N2、H5N6、H5N8等。2020年的秋冬開始,一種H5N1亞型高病原性禽流感病毒在西歐、北美洲與亞洲陸續蔓延,造成許多國家的家禽產業蒙受重大的損失。
禽流感病毒好複雜?不同亞型怎麼區分?
前面提到幾個關於禽流感的名詞,由於與禽流感病毒的特性有關,必須簡要地解釋一下。
所謂禽流感病毒「亞型」,是利用禽流感病毒的兩種蛋白質來區分的:一種蛋白質叫做血球凝集素(hemagglutinin),分成H1至H16共有16亞型(不含國外的蝙蝠發現的H17與H18亞型);另一種蛋白質叫做神經胺酸酶(Neuraminidase),分成N1至N9共有9亞型。以上兩種蛋白質可以組合出16乘以9共144種組合,如H1N2、H5N1、H12N8,這就是我們平常聽到的禽流感病毒亞型。
要特別強調地是,亞型相同的禽流感病毒,致病能力與病毒性狀未必相同,這是因為每一種亞型的蛋白質之間仍有微幅的差別;更何況,禽流感病毒的基因體共有8段,致使自然界中的禽流感病毒性狀非常多樣化。以我國來說,目前在禽場可以發現的H5N2亞型,至少就包含低病原性的美洲株H5N2亞型,和高病原性的2.3.4.4b分支H5N2亞型病毒;這兩者都是H5N2亞型,但病原性差異很大。
「病原性高低」代表對雞隻致病力、致死率
禽流感病毒是否具有「高病原性」,是依據病毒對雞的致病能力來區分的。致病能力可以利用感染實驗雞之後雞隻出現的症狀和死亡率來鑑定,也可以藉由解讀病毒蛋白質特定區域的胺基酸序列判定。因此,高病原性禽流感病毒感染鴨子、鵪鶉或其他鳥類的時候,並不一定會呈現明顯的症狀或很高的死亡率。
目前已知的高病原性禽流感病毒都是H5亞型或H7亞型;但反過來說,H5亞型或H7亞型的病毒不一定就是高病原性的病毒(如前一段舉例的H5N2亞型禽流感病毒),還是要看胺基酸序列或是動物試驗的結果才能判定。
細讀近年有關高病原性禽流感病毒報導的人,可能會留意到有時文章中會冒出「病毒屬於2.3.4.4演化分支」的字眼,但看不懂「演化分支」指的是什麼。文章開頭提到,近二十餘年在全球造成數波大流行的高病原性禽流感,源自於1990年代發現的H5亞型病毒。這一株病毒在自然界中經過長期的演變,衍生出許多有不同差異的子子孫孫,研究禽流感的科學家們為了研究上的方便與促進學者之間的溝通,依據病毒血球凝集素基因的相似程度,將這些子孫們歸類與編碼,於是有了「1.3.2.1」、「2.3.4.4 c」這樣類似章節段落編排的分類系統。
禽流感病毒的傳播方式:人、車與設備都是潛在攜帶者
禽流感病毒的傳播,可概略分為長距離和短距離傳播。
長距離的傳播指的是跨越陸塊、穿越國境的傳播。自然界的禽流感病毒長距離傳播主要依賴候鳥季節性遷移時的攜帶,傳播的距離往往以數千公里計,甚至越過赤道,橫跨南北半球。
再者,候鳥的遷徙與季節改變有著緊密的關聯,但候鳥遷徙未必全是高低緯度之間的南北向移動。如圖一所示的「大西洋東岸線」、「黑海與地中海線」、「東亞澳洲線」區域內的候鳥可以東西向遷徙,在不同的陸塊之間移動。因此,禽流感病毒隨著禽鳥散播也會是東西向的,譬如2014年冬天北美洲爆發的高病原性禽流感,就是從俄羅斯的西伯利亞向東跨越白令海峽進入美國阿拉斯加州,再南下擴散到加拿大和美國本土的。

圖一、全球候鳥的8條主要遷徙路線。由圖左側起:太平洋美洲沿岸線(深綠色圍繞的區域)、美洲縱貫線(淺黃色)、大西洋西岸線(淺藍色)、大西洋東岸線(紅色)、黑海與地中海線(淺綠色)、東亞與東非線(深藍色)、中亞線(粉紅色)、東亞澳洲線(橙黃色)。(圖片引用自「歐亞非候鳥保育協議」(AEWA)網站)
人為的長距離傳播主要是活禽與動物產品的運輸,空運與海運均有可能,所以動物與動物品的跨國運輸需要透過國與國之間的協議與嚴密的檢疫,方能降低疫病傳播的風險。在航空旅行便捷的今天,人類感染後攜帶禽流感病毒長距離傳播當然也有機會。
短距離的傳播是指病毒在一個陸域內的畜牧場之間傳播,距離通常以數十至數百公里為尺度。一如非洲豬瘟與口蹄疫等高傳染性的動物傳染病,禽流感病毒的短距離傳播與人類的行為密切相關。沒有經過充分清洗消毒的運輸車輛、籠具以及未遵守畜牧場生物安全原則出入畜牧場的人員、車輛與設備,都是潛在的病毒攜帶者。相鄰禽場之間的空氣傳播雖然也有可能,但並非禽流感病毒的主要傳播途徑。
- 延伸閱讀:疫情週期可以預測嗎?禽流感小知識大問答
近期H5N1禽流感疫情嚴峻,全球處置、死亡家禽至少2億隻
H5亞型禽流感在21世紀造成全球性的大規模疫情,大致上可以分為三波大流行:2005年至2010年、2011年至2019年、2020年至今(圖二)。
最近的一波疫情始於2020年在歐亞大陸的H5N8亞型疫情,2021年的病毒亞型由H5N8亞型逐漸轉換為H5N1亞型,並且一路延續至今。這一波全球疫情造成各類家禽的大量死亡,因病死亡或人道處置的總數累計至少2億隻,超過前兩波疫情的總和。由於疫情仍在各地持續,數字必將不斷攀升。
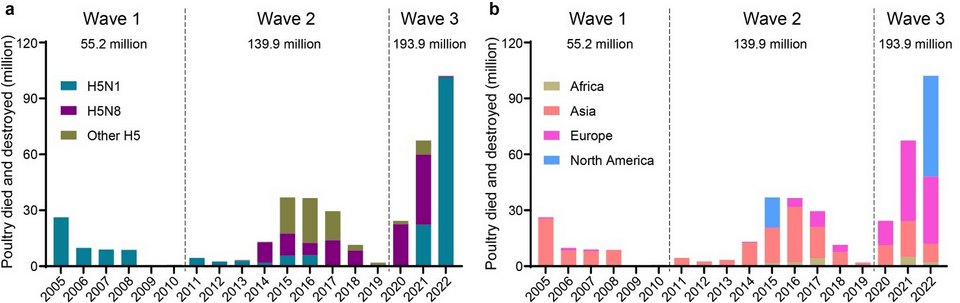
圖二、H5亞型禽流感在21世紀造成的三波全球性大流行。左側圖(a)顯示H5N1(藍綠色)是第一波及最近的第三波的主要病毒亞型。右側圖(b)顯示在三波疫情中,亞洲(橘色)都是主要受災區域;2015年以後歐洲(粉紅色)與北美洲(淺藍色)也出現嚴重疫情。(取自Shi等;Emerging Microbes & Infections, DOI: 10.1080/22221751.2022.2155072)
從近幾個月的數字來看,東亞國家在2022年入秋以後,H5N1亞型感染的家禽場首次出現於11月15日,地點是韓國西北部的京畿道龍仁市;截至年底案例場已有54場,約324萬隻家禽死亡或遭到人道處置,韓國全境除了濟州道以外均有疫情。日本的疫情爆發於11月28日的岡山縣,到2022年底為止,日本的案例場數已累積49場,遍及22個道縣,730萬隻家禽死亡或遭到人道處置。印度、以色列、越南與俄羅斯鄰近太平洋的省份,今年也有H5N1亞型感染家禽場的案例通報。中國大陸除了在野鳥檢出H5N1亞型病毒外,並未通報任何家禽場的案例。
歐洲的疫情依然嚴峻,2022年的第4季有27個國家發生高病原性禽流感,撲殺了大約4千萬隻家禽。第4季共有398個禽場遭到高病原性禽流感病毒入侵,受感染的禽場以英國、法國、匈牙利、德國境內較多。
北美洲近兩年算是禽流感的重災區。美國在2022年有47州爆發H5N1亞型高病原性禽流感,僅有西維吉尼亞州、路易斯安那州、夏威夷州3州倖免。全年共有715個家禽場確診,其中306場為商業化禽場,409場為後院式的小規模禽場,受害的家禽總數逼近5千8百萬隻。經歷了前一個冬天的疫情,2022年歲末的第四季疫情並未趨緩,依據美國農業部公布的數據,2022年10月至12月每月感染家禽總數分別是211萬、375萬、507萬,依然呈現上升的趨勢。加拿大的疫情規模雖然沒有美國大,但2022年一整年也有約300萬隻家禽受到禽流感侵害,受害的範圍達8個省。
另一個不好的消息是,過去H5亞型禽流感在美洲的疫情多半發生在北美洲,但2022年年底部分中南美洲國家也淪為疫區。哥倫比亞與厄瓜多的家禽場爆發了H5N1亞型禽流感案例,委內瑞拉、祕魯、智利等國也有鵜鶘感染的案例。

國內關於國際禽流感疫情報導較少,其實非洲近年也頗受影響,包括野生動物,如去(2022)年瀕危物種非洲企鵝便傳出感染禽流感死亡案例。(圖片來源/kallerna CC BY-SA 4.0)
我國在2021年前並未在野鳥或家禽場發現H5N1亞型高病原性禽流感病毒。第一次在國內田間檢出H5N1亞型病毒,是2022年8月在花蓮縣死亡的黑腹燕鷗與蒙古鴴等野鳥身上,偵測到同一時期流行在東亞地區的H5N1亞型病毒。家禽場的第一個確診案例出現在2022年11月20日,地點是宜蘭縣五結鄉。隨後彰化縣、雲林縣、嘉義縣、臺南市陸續出現案例,感染家禽的種類含括有色肉雞、土番鴨、正番鴨、肉鵝、蛋雞,短短一個多月就有17個禽場感染H5N1亞型病毒。同時期除了新入侵的H5N1亞型病毒,也有既有H5N2亞型高病原性禽流感案例,讓情勢益加複雜。
哺乳動物與人類感染的風險,須持續關注
目前對當下流行的H5N1亞型禽流感病毒的分析,都認為目前的人類感染病例都是偶發的案例,病毒不太容易感染人類,更不容易造成透過人傳人的途徑在人類社會蔓延開來。但是依據世界衛生組織自2003年以來的資料蒐集,H5N1亞型禽流感病毒感染人類的致死率累計達56%(135/240),感染人類後的嚴重程度不可小覷。
另外,隨著H5N1亞型病毒的散播,在歐洲與美洲已經出現感染紅狐、條紋臭鼬、美洲黑熊等哺乳動物的案例。雖然這些案例為數不多,依然顯示這個病毒株可能逐漸增加對哺乳動物的感染能力,必須持續關注它的變化。
如何因應禽流感的全球性疫情?牧場生物安全是首要
禽流感病毒的演化是大自然的一部份,候鳥的遷徙也是生物適應性演化的結果。與其眼看著疫情發生,追究病毒「為何」會變成現在我們看到的樣貌,不如把問題拉回到比較務實的層面。
首先,防堵或干擾候鳥的遷徙,既違反自然界的規律,也不合生態環境保育與永續經營的普世價值。既然天命不可違,或許就該回頭看看人類可以做點什麼。如果把候鳥和病毒想像成颱風之類的天災,進入雨季之前,是不是應該檢修房舍和疏通溝渠?颱風接近、警報響起之時,是不是應該儲備糧食、迅速避難以尋求減災?把相同的概念套用在禽流感的防疫上,就是「做好牧場生物安全」的概念。
「牧場生物安全」看似老生常談,其實最為實用。登記牧場每日的家禽死亡,掌握家禽細微的健康與行為變化;檢修房舍與鳥網,防杜獸禽蟲鼠自由進出禽舍;做好人員和車輛進出管制,工作區與一般區的嚴謹區劃,避免病原進入禽舍;落實人員訓練,建立牧場內清潔消毒的好文化;遇到健康異常趕緊通報獸醫師,讓專業協助把關。魔鬼總是藏在細節裡,這些生物安全工作若能落實,就算沒有辦法把牧場感染禽流感的風險降到零,也已經盡力將風險最小化。更何況牧場生物安全對抗的不僅是禽流感,而是對場內動物健康的整體提升,效益是全面性的。

2017年喀麥隆爆發H5N1禽流感後,當地雞農身穿防護服去為養雞隻。(圖片來源/IAEA Imagebank CC BY-NC-ND 2.0)
其次,想要單純依賴分子生物學技術當作疫情調查、「快速」追溯疫情的主要工具或許並不實際。禽流感病毒的基因體有8段,快速診斷僅檢測血球凝集素基因上一個長度為數百個核苷酸的小區域並進行序列解讀,打算利用1、2個核苷酸的微幅差異解讀感染場之間是否有疫情相關性,實在是高估了分子流行病學的可能性。
基因體全長序列分析雖然在擁有次世代定序技術的今天已經能更有效率的執行,可是投入的經費不低,時間上也依然追不上疫情的發展;能否獲得完整的序列更要視檢體的種類與品質而定。疫情調查還有很多面向,詳實地了解禽場的人車出入與日常活動紀錄,可能更加具體且容易找到改善的方向。以這3年的新型冠狀病毒感染為例,防範社區群聚感染,不也是仰賴實名制與出入特定場所的紀錄?
疫病、戰爭等各方面影響家禽產業,反求諸己才能永續經營
關於前年以來的雞蛋供給,能不能歸咎於這波禽流感疫情,其實頗難論斷。從全球的大尺度來看,因為同時間人類社會正在奮力與新冠病毒相搏,人禍又有俄烏戰爭,包括畜產業在內的各行各業無不窮於應對,從上游飼料原料供給到下游運輸、食品加工的人力缺口,已經很難找到單一因子去解釋供應鏈的全面失序。
即便只看臺灣單一國家,雞蛋的供需與價格漲跌也是多因子互動下的產物,無法讓禽流感背負全部的責任。何況國內民眾對動物傳染病主動通報尚未蔚為風氣,確診數字與實際感染場數的落差不明,所以想從僅有的確診案例數字推論禽流感與蛋價的關係,無異以管窺天、盲人摸象,需要更多、更具代表性的數據才有討論的空間。
總結來說,從趨勢上來看,進入21世紀以來H5亞型禽流感的疫情起起伏伏,H5N1亞型近2年的肆虐只是另一個新角色的登場,朝代更迭但劇本基本上沒有太大的改變。無論禽流感的疫情與全球暖化、氣候變遷能不能掛上關聯,病毒的轉換一如候鳥的春去秋來,或許也可視為一種新的常態。若是如此,站在與地球永續共存的角度,反求諸己,鞏固牧場生物安全這把防疫的保護傘,才能讓產業永續。
延伸閱讀:
- 疫情週期可以預測嗎?禽流感小知識大問答
- 金門檢出國內首例「可禽傳人」H9N2亞型低病原禽流感 疾管署:人類致死率低
- 雞蛋風暴不再來 蛋雞產業結構調整不斷鏈 蛋中雞場將加強管理
- H5N1虐全球 重啟禽流感疫苗討論 改採載體疫苗有機會降低病毒變異風險